
A maioria dos inseticidas comercialmente importantes para a produtividade dos nossos agrossistemas são neurotoxinas que atuam dentro do sistema nervoso dos insetos. Contudo, não é de se espantar que ainda tenhamos diversos questionamentos sobre estes inseticidas. Por exemplo: Compostos vegetais possuem atividade inseticida? Inseticidas de ocorrência natural coevoluíram com o seu organismo? Qual é seu modo e sítio de ação? Como ocorre a resistência a um determinado inseticida? E, por que se proteger com equipamento de proteção individual (EPI)?
É por isso que este artigo foi concebido, justamente para responder, de forma sucinta, a todas estas perguntas.
Alcaloides vegetais – agonista (compostos que se ligam aos receptores de neurotransmissores para estimular a abertura dos canais iônicos) ou antagonista (compostos que se ligam aos receptores de neurotransmissores para fechar canais iônicos) – são compostos que podem apresentar propriedades inseticidas. Por exemplo, a nicotina (Figura 1), um bioinseticida de ocorrência natural que vem sendo bastante empregado para o controle de pragas agrícolas há centenas de anos, é um alcaloide de planta de tabaco (gênero Nicotiana), que tem o mesmo modus operandi da acetilcolina (Figura 1), um dos principais neurotransmissor do cérebro do inseto; contudo, este alcaloide é altamente tóxico a outros organismo não-alvo, como os mamíferos. Outros exemplos de alcaloides, com propriedades inseticidas, incluem anabasina (do tabaco), a citisina (do Laburnum), dihidro-β-erythroidina (da Erythrina), lobelina (da Lobelia), metilicaconitina (da Larkspur) e tubocurarina (da Chondrodendron). Ao que parece, estas substâncias coevoluíram com suas plantas como um mecanismo de proteção contra insetos e outros herbívoros; estes compostos, no entanto, não têm sido utilizados comercialmente – como bioinseticidas – devido à baixa seletividade a espécies não-alvo.
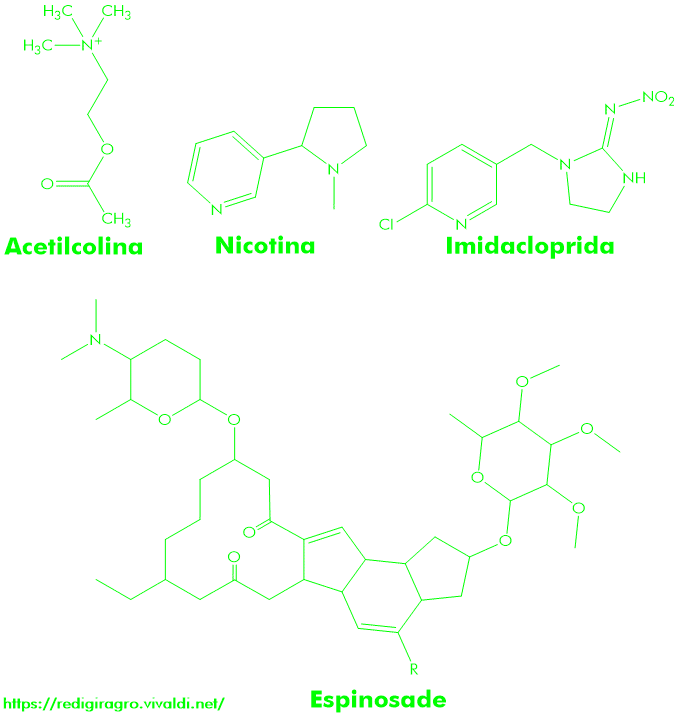
Figura 1: Estrutura química da acetilcolina (um dos principais – e mais rápido – neurotransmissor dos insetos), da nicotina (um bioinseticida de ocorrência natural isolado da planta do tabaco Nicotiana, que possui atividade inseticida nos receptores nicotínicos), da imidacloprida (primeiro ingrediente ativo do grupo dos neonicotinoides), e da espinosade (mistura de duas lactonas macrocíclicas, isoladas do actinomiceto Saccharopolyspora spinosa, uma bactéria do solo, fixadora de dinitrogênio atmosférico (N2).
Agora que já sabemos que inseticidas são neurotoxinas, resta-nos saber o que são neurotoxinas e como agem, e é o que veremos a seguir.
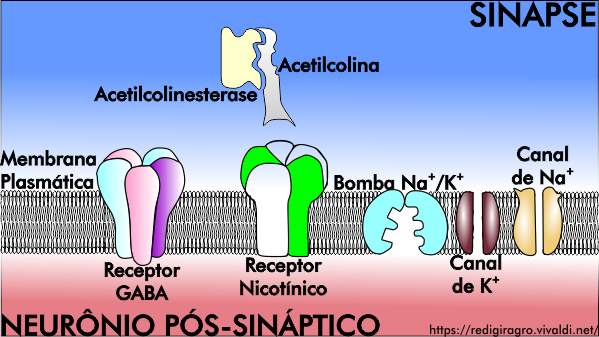
Neurotoxinas são substâncias que intoxicam o sistema neuromuscular dos insetos: por exemplo, os carbamatos e organofosforados são inibidores da enzima acetilcolinesterase, os ciclodienos e fenilpirazóis são bloqueadores dos canais de cloro, os piretroides e DDTs retardam o fechamento dos canais sódio dependentes do potencial de membrana e, neonicotinoides e espinosinas imitam e competem com acetilcona pelo sítio de ação nos receptores nicotínicos. Portanto, o sistema neuromuscular dos insetos é composto por receptores nicotínicos (neurais e musculares), receptores do neurotransmissor ácido γ-aminobutírico (ou GABA), acetilcolina e acetilcolinesterase, bomba de sódio e potássio, canais iônicos, entre outros componentes (Figura 1). Mas como agem as neurotoxinas no sistema neuromuscular?
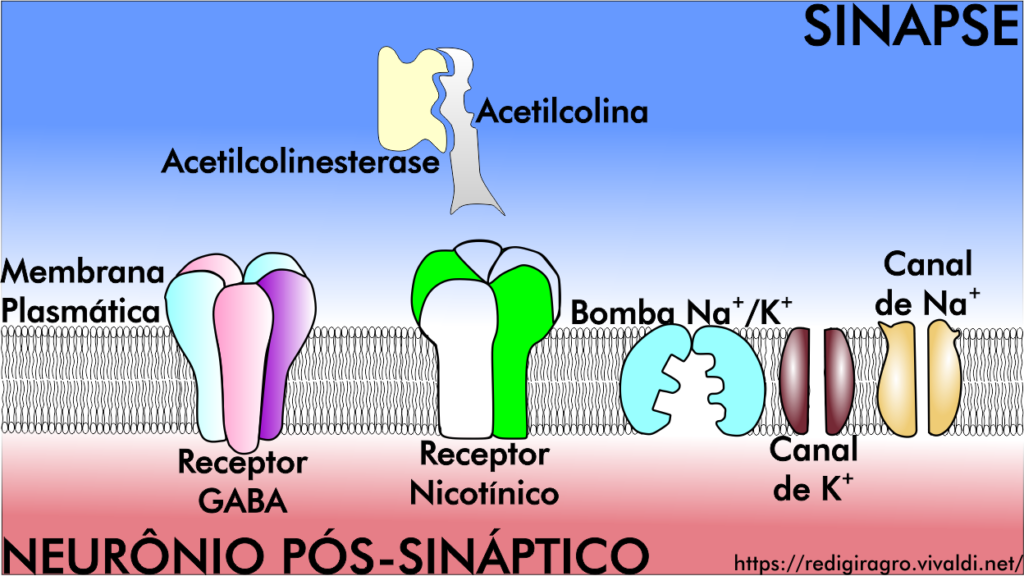
Figura 2: Componentes do sistema neuromuscular dos insetos.
Os neonicotinoides e espinosinas são o grupo de inseticidas neurotóxicos de maior importância para proteção da saúde vegetal e animal, uma vez que atuam de forma complementar, ou seja, agem em diferentes grupos de receptores nicotínicos, proporcionando um amplo espectro de controle a pragas agrícolas associado à baixa toxidade a mamíferos. Receptores nicotínicos, localizados na superfície da membrana plasmática de células nervosas (Figura 2), são membros de uma superfamília de canais iônicos dependentes de ligantes, e são o sítio de ação de uma plêiade de compostos sintéticos ou de ocorrência natural que apresentam atividade inseticida; são assim chamados, porque só se descobriu que acetilcolina os ativava utilizando a nicotina como agonista.
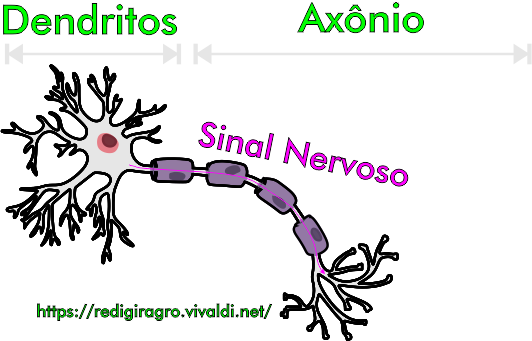
Figura 3: Estrutura de uma célula nervoso (ou neurônio), ilustrando o sentido do sinal nervoso (ou impulso nervoso).
Os neonicotinoides (Figura 1), moléculas sintéticas análogas à nicotina, após serem absorvidos pelos insetos, se ligam aos receptores nicotínicos e ativam o influxo de sódio (de carga positiva) no terminal pós-sináptico (ou seja, nos dendritos), o qual é o responsável pela continuidade do impulso nervoso (elétrico) recebido, nos axônios pré-sinápticos, pelas vesículas que transportam acetilcolina, as quais, após serem estimuladas por este impulso, passam a se acoplar à membrana plasmática do neurônio pré-sináptico (Figura 2, Figura 3. Não deixe de ver a animação!), para liberar acetilcolina na sinapse (região que possibilita a comunicação entre um axônio e dendritos de células nervosas vizinhas). A interrupção da transmissão do sinal nervoso (ou seja, do influxo de sódio, de carga positiva) através da membrana do neurônio pós-sináptico é feita pela acetilcolinesterase, que pode degradar até 25 mil moléculas de acetilcolina (em ácido acético e colina) por segundo, evitando assim que elas se liguem aos receptores nicotínicos e ativem-nos. Contudo, a acetilcolinesterase não tem o mesmo efeito sobre nicotina e neonicotinoides, o que significa dizer que, quando estes dois compostos se ligam aos receptores nicotínicos, haverá um influxo de sódio contínuo que levará à despolarização da membrana plasmática e à hiperexcitação do sistema nervoso, podendo, no final, ocasionar a morte do inseto; a imidacloprida, primeiro ingrediente ativo do grupo de inseticidas neonicotinoide, apresenta forte atividade inseticida sobre fitossugadores da ordem Hemiptera, tais como pulgões, moscas-brancas e cigarrinhas, e sobre alguns coleópteros, tais como besouro-da-batata (Leptinotarsa Decemlineata), assim como contra pulgas de gatos (Ctenocephalides felis) e de cães (Ctenocephalides canis).
As espinosades (Figura 1), uma mistura de duas lactonas macrocíclicas, espinosade A e D, são ingredientes ativos de ocorrência natural, do grupo das espinosinas, que foram inicialmente isoladas do actinomiceto Saccharopolyspora spinosa, uma bactéria do solo, que fixa N2 (dinitrogênio atmosférico); estas neurotoxinas são eficaz para o controle de pragas como lagartas (Lepidoptera), tripes (Thysanoptera) e moscas (Diptera), pois ativam, nos neurônios pós-sinápticos, os receptores nicotínicos; vale ressaltar ainda que o sítio de ação das espinosinas é diferente do sítio de ação dos neonicotinoides: por exemplo, na mosca-da-fruta (Drosophila melanogaster), as espinosinas atuam na subunidade Dα6 e os neonicotinoides, nas subunidades Dα1, Dα2, Dα3.
Carbamatos e organofosforados são mais precisos do que neonicotinoides e espinosinas, pois se ligam diretamente à acetilcolinesterase e inativam-na.
Piretroides e DDTs retardam o fechamento de canais de sódio dependentes do potencial de membrana após a transmissão do potencial elétrico pela membrana plasmática do neurônio pós-sináptico (em direção às sinapses), o que leva a um grande dispêndio de energia na forma de ATP, uma vez que o sistema nervoso dos animais gasta cerca de 60% de sua energia, ou seja, 30% de toda energia do animal, para bombear sódio para as sinapses, por meio da bomba sódio/potássio, o que evidentemente comprometerá outras funções vitais do inseto levando-o, portanto, à morte. Importante ressaltar que os piretroides são moléculas análogas às piretrinas que podem ser extraídas de flores de crisântemo.
Ciclodienos e fenilpirazóis, por outro lado, são bloqueadores dos canais de cloro dependentes do ácido γ-aminobutírico (GABA). Em condições normais, estes canais de cloro são ativados pelo neurotransmissor ácido γ-aminobutírico quando este se liga ao receptor GABA, o que aciona o influxo do cloro (de carga negativa), cuja função é a inibição da transmissão do sinal nervoso (ou do fluxo de sódio, cujo sinal é positivo) pelo neurônio pós-sináptico.
De forma geral, podemos, sim, dizer que as neurotoxinas ativam canais de sódio dos insetos, despolarizam suas membranas plasmáticas (através do influxo de sódio), ocasionam dispêndio de energia e a hiperexcitação do sistema neuromuscular, o que, em última instância, leva a tremores musculares, a paralisia, a convulsões e, por fim, eventualmente à morte do inseto.
Ora, e se nós humanos também possuímos receptores nicotínicos, como devemos nos comportar face a estes inseticidas?
Devemos nos proteger destes inseticidas usando equipamento de proteção individual (roupa impermeável, botas, óculos de proteção e/ou protetor facial, e luvas) , pois sabemos que nossos receptores nicotínicos neurais medeiam efeitos da nicotina associados ao tabagismo, e que, para além disso, estes receptores estão associados com desordens neurológicas, tais como déficit de atenção, Alzhemeier, epilepsia, Parkinson, esquizofrenia e síndrome de Tourette; por outro lado, o mal funcionamento dos receptores nicotínicos musculares ocasiona desordens neuromusculares, tais como miastenia grave e miastenia congênita. Portanto, uso do equipamento de proteção individual é imprescindível para preservação da saúde do aplicador (do inseticida).
Para finalizar este artigo, vamos entrar no âmbito da resistência à inseticida, ou seja, como o inseto se protege contra inseticidas? Os insetos podem desenvolver resistência à inseticidas por aumento da detoxificação (por meio da elevação da atividade metabólica e/ou pelo aumento de enzimas sequestradoras de inseticidas) e/ou por alteração estrutural do sítio de ação do inseticida. Como ocorre a resistência por alteração do sítio de ação?
Para entendermos isso, precisamos primeiro entender os componentes dos receptores nicotínicos.
Receptores nicotínicos são, em geral, pentâmeros, isto é, composto de 5 subunidades que podem ser α e não-α (β, γ). Cada subunidade tem 4 domínios transmenbrana e um enorme domínio extracelular N-terminal – composto de aproximadamente 200 aminoácidos. Receptores nicotínicos da mosca-da-fruta (Drosophila melanogaster), um inseto-modelo, contudo, possuem 10 subunidades (Dα1 – Dα7 e Dβ1 – Dβ3)). Estas subunidades podem se rearranjar de modo a formar novas populações de receptores nicotínicos, e este rearranjo facilita desenvolvimento da resistência contra inseticidas.
Para falar da alteração do sítio de ação, usarei como exemplo a orizicultura asiática: na Ásia, uma das principais pragas do arroz, a cigarrinha-marrom (Nilaparvata lugens), desenvolveu 250 vezes mais resistência a imidacloprida (um neonicotinoide) por alteração estrutural do sítio de ação; na verdade, esta praga apenas trocou a tirosina (um aminoácido), da posição 151 do sítio de ligação a neonicotinoide, por serina em duas subunidades dos seus receptores nicotínicos (N1α1 e N1α3), gerando a mutação Y151S (Y é tirosina; S é serina). O fato mais interessante é que a mutação Y151S não afetou a atividade da acetilcolina, mas provocou uma drástica redução na atividade da imidacloprida e de outros inseticidas comercialmente disponíveis. A resistência a neonicotinoides também tem sido relada em mosca-branca e no besouro-da-batata. Há também relatos da resistência de pulgões à nicotina e das lagartas Spodoptera exigua e Helicoverpa armigera à espinosade.
Agora que você já leu sobre inseticidas que atuam no sistema nervoso, que tal produzir seu próprio inseticida? Para isso, clique aqui.
Caso você necessite aplicar qualquer grupo de inseticidas, incluindo os que estudamos aqui, consulte ingredientes ativos autorizados pelo Ministério da Agricultura, Pecuária e Abastecimento (MAPA) clicando aqui.
Se quiser que eu lhe explique este conteúdo pessoalmente, clique aqui.
Autor: Dr. Carlos Vergara (Ciência do Solo)
Revisor: Dra. Karla Emanuelle Campos Araujo (Professora de Métodos de Controle de Pragas na Faculdade de Ciências Agrárias de Araripina)
Citação: VERGARA. C. Neurotoxinas em pragas agrícolas. Redigiragro (https://redigiragro.vivaldi.net), 2021.
Literatura
MILLAR, Neil S.; DENHOLM, Ian. Nicotinic acetylcholine receptors: targets for commercially important insecticides. Invertebrate Neuroscience, 2007, 7.1: 53-66.
MALONEY, E. M., et al. Binding properties to nicotinic acetylcholine receptors can explain differential toxicity of neonicotinoid insecticides in Chironomidae. Aquatic Toxicology, 2021, 230: 105701.
POZEBON, H.; ARNEMANN, J. A. Como funcionam os inseticidas carbamatos e organofosforados? Portal Mais Soja. 2020. Disponível online.